INTRODUCCIÓN
Se puede hablar de la respiración como un proceso esencial para la vida. A excepción de algunos microorganismos, todos los seres vivos respiran, de una forma u otra. No obstante, la respiración puede verse interrumpida por múltiples factores, pudiendo provocar la muerte. Por suerte, los avances en medicina y, específicamente, en la resucitación han hecho que el cese de la respiración no siempre preceda a un desenlace desafortunado.
Ya en la mitología griega Isis, diosa del misterio y la sabiduría, intenta resucitar a Osiris introduciendo el aire que nace del movimiento de sus alas hacia el interior de su cuerpo, ésta es la primera referencia que se conoce respecto a la intención de resucitación de una persona a través de la insuflación de aire.
De igual forma Galeno dejó reflejado en su libro Procedimientos de anatomía (año 175 d.C.) la importancia de la respiración artificial para evitar el colapso de los pulmones cuando realizaba las toracotomías a los animales.
Años después fue Paracelso (1493-1541 d.C) quien experimentaba la reanimación a un paciente colocando un tubo en su boca e insuflándole aire mediante un fuelle. Vesalio (1543 d.C) hará lo propio realizando una apertura en la tráquea y colocando un tubo de junco o mimbre desde donde se insuflaría el aire.
En 1754, Black, Priesltley, Lavoisier y Scheele publicaron su descubrimiento sobre los gases implicados en la respiración, el dióxido de carbono y el oxígeno, fundamentos que impulsaron a la construcción de los primeros artilugios de ventilación mecánica o respiradores.
El primer dispositivo de ventilación a presión positiva lo creó Dräger en 1911, al que denominaron Pulmotor. Éste funcionaba mediante un cilindro de oxígeno o aire comprimido como fuente de energía y dotaba al paciente, mediante una mascarilla nasobucal, de una mezcla de gases y aire ambiente.

Imagen 1: El Pulmotor. Fuente: Zivilschutzmuseum Krankenbett.JPG
Unos años más tarde, en 1927, el ingeniero estadounidense P. Dinker presentó el pulmón de acero, creado para usarse en personas con lesiones en la pared muscular del tórax que impidieran la respiración. Éste consistía en un bidón cilíndrico de acero donde se introducía todo el cuerpo de la persona a excepción de la cabeza, ejerciendo en el tórax, y de forma intermitente, presión negativa mediante el bombeo de aire, de esta manera se conseguía una alternanza de la presión intratorácica, y con esto una entrada y salida de aire en los pulmones, imitando así los movimientos respiratorios. Se puede decir que fue la primera forma de ventilación mecánica no invasiva.
El pulmón de acero fue mejorado por la nueva versión más fiable, ligera y económica de J.H. Emerson en 1931. Éste además contaba con velocidades variables de la ventilación y se podía utilizar de forma manual en caso de fallo eléctrico. Fue vital durante la epidemia de poliomielitis que asoló América del Norte y Europa entre los años 1940 y 1950.

Imagen 2: Sala de pulmones de acero llenas con pacientes de poliomielitis. Fuente: http://www.fda.gov/cber/summaries/cent092302pp.htm
Pero mientras los estadounidenses y los ingleses se centraban en la ventilación mediante presión negativa, los daneses, también motivados por la epidemia de poliomielitis que arrasó Dinamarca en 1953, se decantaron por la ventilación con presión positiva. Engström diseñó un nuevo ventilador capaz de regular la cantidad de volumen de aire que se insuflaba al paciente a través de una traqueotomía. Posteriormente, en 1960, aparecerán los respiradores ciclados por presión y por volumen y, en los años setenta, los nuevos modos ventilatorios y la presión positiva al final de la espiración (PEEP), todo ello de suma importancia para la evolución de la ventilación mecánica.
Las salas repletas de pacientes ventilados con presión positiva son la antesala de lo que hoy conocemos como Unidad de Cuidados Intensivos Respiratorios.
La función que asume el sistema respiratorio es principalmente el de intercambio de gases entre oxígeno y dióxido de carbono mediante la respiración, pero también está implicado en un complejo entramado de funciones relacionadas con la homeostasia que repercute a otros sistemas. Si el pulmón no responde adecuadamente y no se actúa en tiempo y forma adecuados, es probable que provoque el fallo de otros órganos.
Para entender las implicaciones de las patologías respiratorias, las respuestas de nuestro organismo y los mecanismos de los procedimientos para combatirlos es importante hacer un breve recuerdo de la anatomía y fisiología de nuestro sistema respiratorio.
1. ANATOMÍA DEL SISTEMA RESPIRATORIO
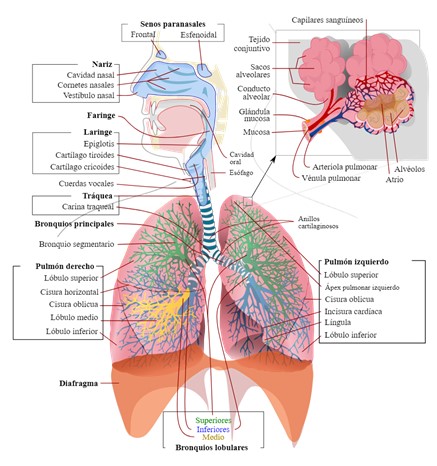
Imagen 3: Esquema del sistema respiratorio. Fuente: Respiratory_system_complete_en.svg
La estructura del sistema respiratorio se divide en dos partes, la vía aérea superior y la vía aérea inferior considerando como hito anatómico el cartílago cricoides. Cada una de estas partes posee unas características concretas para cumplir con su función. A excepción de los alvéolos, el resto de las estructuras son vías aéreas de conducción, es decir, van a dirigir el aire inspirado hasta los alvéolos donde tendrá lugar el intercambio de gases y a canalizarlo de nuevo hacia el exterior mediante la exhalación.
1.1 Vía aérea superior
La vía aérea superior, situada en cabeza y cuello, se compone por la cavidad nasal, la faringe y la laringe.
Esta estructura tiene las características anatómicas idóneas para cumplir la función de protección, especialmente la cavidad nasal que debido a su ángulo de 90º respecto a la tráquea y la presencia de los cornetes permiten concentrar el aire en una corriente pequeña permitiendo calentar, humidificar y filtrar el aire que se inhala. En la nariz se efectuará el primer filtrado a través del vello nasal. A su vez, todas las estructuras del árbol respiratorio están recubierto por un manto mucoso, capaz de atrapar las impurezas del aire y, mediante el movimiento de los cilios de las células epiteliales de la mucosa respiratoria, se evacúan las sustancias irritantes hacia la faringe desde donde podrán ser expulsadas mediante la tos.
La faringe, estructura formada por músculo constrictor, comprende desde la parte posterior de la cavidad nasal hasta la laringe. Se divide en tres partes anatómicas: la nasofaringe, la orofaringe y la laringofaringe.
La primera división anatómica, la nasofaringe, se comunica lateralmente con el oído medio mediante las Trompas de Eustaquio. La orofaringe, también denominada bucofaringe, se limita desde el velo del paladar hasta la epiglotis, y podemos encontrar las amígdalas palatinas (o anginas). Por último, la laringofaringe, también hipofaringe o laringe inferior, comprende desde la epiglotis hasta la entrada de la laringe, que coincidirá dorsalmente con la 6ª vétebra cervical.
En la mucosa faríngea encontramos gran cantidad de vasos linfáticos, muchos de ellos se comunican con las amígdalas, cumpliendo así la función de prevención de infecciones. Por otra parte, la faringe también cumple con la función de discriminar hacia dónde debe dirigir el contenido de la cavidad bucal, es decir, si se trata de un alimento lo dirigirá hacia el esófago, si se trata de una bocanada de aire hacia la laringe. Para ello, la faringe está inervada por las ramas del plexo faríngeo del simpático y las fibras del nervio vago, del parasimpático, que se sincronizan para coordinar cada uno de los movimientos correspondientes de la respiración con los de la deglución. Para ello encontramos, en su unión con la laringe, la epiglotis, una membrana cartilaginosa que se mantendrá abierta para permitir el paso del aire durante la respiración, y se cerrará con el paso de los alimentos facilitando el tránsito correcto hacia el esófago.
La laringe, además de ser una vía de comunicación con las vías respiratorias inferiores, es el órgano encargado de la fonación. Formada por el hueso hioides y por los cartílagos tiroides, cricoides, aritenoides y corniculado, unidos por ligamentos y fuertes músculos. Tapizada internamente por un manto mucoso ciliado, continúa con la función de purificación y humidificación del aire inspirado. En su porción superior protruyen lateralmente dos pliegues de esta capa mucosa hacia la luz, conocidas con el nombre de cuerdas vocales vestibulares o falsas, e inferior a ellas, en la zona denominada glotis, encontramos dos nuevas protusiones a las que se conocen como cuerdas vocales verdaderas. Éstas últimas son las responsables de la emisión del sonido, la fonación, cuando el aire espirado pasa por la glotis y este se estrecha haciendo vibrar las cuerdas vocales.
1.2 Vía aérea inferior
La vía aérea inferior está compuesta por la tráquea, los bronquios y bronquiolos, y los pulmones.
La tráquea es un tubo fibromuscular de entre 10-12 cm de longitud y 16-18 mm de diámetro, formada por anillos de cartílago en forma de “C” incompletos en su parte posterior, los anillos traqueales, que se unen posteriormente a la pared membranosa de la tráquea, otorgando así rigidez estructural a la vía aérea.
La tráquea se bifurca a la altura de la 4ª o 5ª vértebra torácica en dos ramas laterales, los bronquios. La rama derecha con una inclinación de 20°, y la rama izquierda con un eje inclinado de entre 40-50°. Los bronquios siguen la misma estructura de la tráquea de tubo formado por anillos cartilaginosos, el derecho de 2 cm de longitud y de 15 mm de diámetro, el izquierdo de 5 cm de longitud y 11 mm de diámetro. Esta diferencia es debida al espacio ocupado por el corazón.
A continuación, los bronquios se ramifican formando un árbol, las ramas secundarias o bronquios intrapulmonares finalizan en los bronquiolos terminales, último tramo de la vía aérea de conducción que precede a los alvéolos.
El epitelio de la tráquea y los bronquios está compuesto por una capa mucosa que abunda en glándulas mucíparas y células epiteliales ciliadas. Las glándulas mucíparas son las productoras de mucina, que constituye el mucus de la vía aérea, mientras que las células epiteliales ciliadas ayudan a expulsar el mucus hacia el exterior. En los bronquiolos la capa de epitelio cambia a una de células epiteliales no ciliadas que secretan surfactante, lípidos, glicoproteínas y moduladores inflamatorios.
El árbol traqueobronquial está envuelto de músculo liso inervado bajo el control involuntario del plexo pulmonar y del vago.
Protegidos por la caja torácica encontramos los pulmones, dos estructuras de forma cónica con su ápex en la parte superior y la base, o cara diafragmática, en la parte inferior. El pulmón derecho, más voluminoso, se divide por las cisuras en tres lóbulos (superior, medio e inferior), mientras que el izquierdo consta de dos (superior e inferior).
La caja torácica está formada por las vértebras torácicas, las costillas, el esternón y el cartílago costal. No es una estructura estática, sino que el engranaje de las diferentes estructuras permite el movimiento de expansión lateral y anteroposterior de la parrilla costal, fundamental para admitir el aumento de volumen pulmonar durante la inspiración.
Ambos pulmones están separados por el mediastino, un espacio comprendido entre el esternón y la columna vertebral que aloja el corazón, el timo, la tráquea, los bronquios, el esófago y los vasos sanguíneos mayores.
Los pulmones están envueltos por la pleura, una membrana serosa formada por dos hojas, la pleura visceral (en unión con la superficie pulmonar) y la pleura parietal (en contacto con la caja torácica y el diafragma). Entre ambas pleuras encontramos la cavidad pleural, espacio que contiene el líquido pleural.
El líquido pleural, con un volumen mínimo de 1.2 ml entre pleuras, tiene la función de mantener ambas membranas unidas, aunque manteniendo la capacidad de deslizarse suavemente la una sobre la otra durante la dinámica respiratoria.
Una vez dentro de la estructura pulmonar, los bronquiolos terminales se ramifican en los conductos alveolares que finalizarán en los sacos alveolares, constituidos por numerosos alvéolos envueltos por una tupida red de capilares sanguíneos.
Se estima que el pulmón tiene entre 300 y 480 millones de alvéolos, con una superficie alveolar útil para el intercambio gaseoso equivalente a unas 40 veces la superficie externa de nuestro cuerpo.
La pared alveolar está formada por dos tipos de células. El 95% de la superficie alveolar son neumocitos tipo I, células planas que forman un epitelio escamoso de una única capa muy fina permitiendo así el continuo paso de los gases. El otro 5 % está compuesto por los neumocitos tipo II, células cuboidales dispuestas de microvellosidades en su cara interior, encargadas de la producción del surfactante, una lipoproteína de acción tensoactiva que se segrega hacia el interior del alveolo disminuyendo la tensión superficial en la interfase alveolar aire-líquido evitando de esa manera el colapso del alveolo.
Los alvéolos además de estar rodeados por la red capilar, también lo está de fibras elásticas, con la función de mantener unidos los alvéolos y de expandirlos durante la inspiración y contraer de nuevo en la espiración. Con ello se consigue que la espiración sea un movimiento pasivo en el que no hay gasto de energía.
1.3. Los musculos respiratorios
Los músculos de tórax y abdomen ejercen un papel protagonista en la mecánica respiratoria.
Los músculos que intervienen en la mecánica respiratoria se juntan en cuatro grupos: el diafragma, los intercostales, los abdominales y los músculos accesorios.
El diafragma es considerado el principal músculo respiratorio. Es un músculo plano que separa la cavidad torácica de la abdominal. Localizado debajo de los pulmones, se inserta por la cara anterior en el apéndice xifoides y por su parte posterior en la columna vertebral. Por él atraviesan varios orificios por donde pasan los vasos principales y el esófago.
El diafragma está compuesto por tres tipos de fibras musculares diferenciándose por sus propiedades fisiológicas e histoquímicas. El 25% de las fibras que constituyen el diafragma son las fibras oxidativas de contracción lenta (FO) con capacidad de producir un bajo nivel de fuerza y muy resistentes a la fatiga. Otro 25% del músculo está constituido por fibras oxidativas glucolíticas (FOG), con un alto nivel de fuerza y relativa resistencia a la fatiga. Y las más abundantes, con un 50%, son las fibras glucolíticas de contracción rápida (FG), responsables del mayor nivel de fuerza y más susceptibles a la fatiga.
La inervación del diafragma la realiza mayoritariamente los nervios frénicos. Cada nervio frénico inerva una hemidiafragma, por lo que cada cavidad pulmonar puede funcionar de manera independiente.
Los músculos intercostales incluyen los intercostales internos y los intercostales externos. Son láminas musculares ubicadas en la parte lateral y anterior del tórax y que ocupan los espacios comprendidos entre dos costillas paralelas, con diferentes longitud y dirección de fibras. Los músculos intercostales están inervados por los nervios espinales torácicos.
En el grupo de músculos abdominales se incluyen los rectos, los transversos del abdomen y los oblicuos internos y externos.
Por último, los músculos accesorios que, aunque no son los principales responsables del movimiento en la respiración sí que otorgan asistencia a éste, son los escalenos, los esternocleidomastoideos, los trapezoides, los pectorales mayor y menor, el serrato anterior y el dorsal ancho.
1.4. Inervación del sistema respiratorio
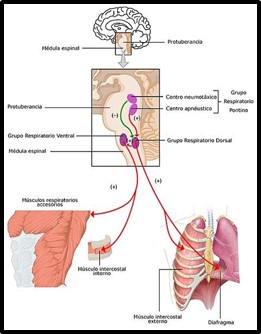
Imagen 4: Centros respiratorios del cerebro. Fuente: 2327_Respiratory_Centers_of_the_Brain_esp.jpg
La retroalimentación que ocurre entre diversos receptores químicos o mecánicos con el sistema nervioso central determina la regulación de la respiración estimulando a sus efectores, los músculos respiratorios.
Los quimiorreceptores, regidos por los pares craneales IX y X, pueden ser centrales o periféricos. Los receptores centrales, ubicados en el bulbo raquídeo y troncoencéfalo, principalmente, son sensibles a los cambios de CO2. Mientras que los quimiorreceptores periféricos ubicados en los cuerpos carotídeos y aórticos son más sensibles a la hipoxemia (O2) y a los cambios de pH.
Los mecanorreceptores pueden ser de distensión, de irritación y los yuxtacapilares. Los receptores de distensión, ubicados en el músculo liso de la vía aérea, son de adaptación lenta y se activan con la elongación de las fibras en la inspiración. Los receptores de la irritación, ubicados en la laringe y en el resto de la vía aérea, son de respuesta rápida mediante la tos, la constricción laríngea, la polipnea o la broncoconstricción. Se estimulan con los gases irritantes, estímulos mecánicos, histamina, reacciones alérgicas, etc. Por último, los receptores yuxtacapilares se encuentran ubicados en el intersticio alveolar y son estimulados con el edema alveolar, o la acción de irritantes químicos. Su respuesta es a través del nervio vago produciendo taquipnea y sensación de disnea.
1.5. Circulación del sistema pulmonar
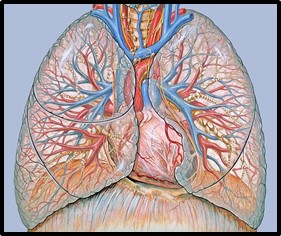
Imagen 5: Circulación pulmonar. Fuente: Anatomía torácica.jpg
La circulación del sistema respiratorio presenta dos ocupaciones bien diferenciadas: el intercambio de gases y la nutrición del sistema.
La circulación pulmonar o menor posee el desempeño funcional del intercambio de gases entre la sangre y el aire. Para conseguirlo las dos ramas de la arteria pulmonar entran en los pulmones a nivel del hilio, en paralelo a la ramificación bronquial éstas se van dividiendo hasta convertirse en las arteriolas alveolares formando una espesa red capilar que rodea el alveolo. Confluyen en las vénulas, discurriendo entre los septos interlobulares y uniéndose en ramas venosas hasta constituir las dos venas pulmonares que salen por el hilio pulmonar hasta llegar a la aurícula izquierda del corazón.
El tiempo circulatorio total por la circulación pulmonar es de 4-5 segundos.
La delgadez de las paredes de sus vasos y la escasez de músculo liso proporcionan una distensibilidad elevada, haciéndolo especialmente susceptible a la deformación por fuerzas externas.
Además, los vasos pulmonares presentan otras funciones muy interesantes para el organismo: liberan sustancias fibrinolíticas protegiendo al cuerpo contra los pequeños trombos, absorbe a través de las paredes de los capilares pulmonares los pequeños émbolos de aire, metaboliza hormonas vasoactivas como la angiotensina II o inactivan otras como la serotonina y participa en mecanismos de defensa celular y humoral pulmonar a través de las inmunoglobulinas (IgA) y de los macrófagos pulmonares.
La circulación bronquial o mayor es la encargada de nutrir las estructuras celulares de los pulmones y bronquios. Formada por las arterias bronquiales que se ramifican desde la aorta y se van dividiendo por la pared de los bronquios hasta formar dos redes capilares, una profunda que nutrirá los músculos y las glándulas, y otra superficial para la mucosa. A continuación, confluirán hasta formar las venas bronquiales que salen por el hilio y desembocan en las venas ácigos y hemiácigos.
El volumen sanguíneo necesario para la circulación pulmonar es de 450-500 ml, representando un 9-10% del volumen total de sangre del organismo y más del 40% del peso del pulmón.
2. FISIOLOGÍA DEL SISTEMA RESPIRATORIO.
La principal función del sistema respiratorio es poner a disposición del organismo el oxígeno (O2) procedente de la atmósfera y expulsar el dióxido de carbono (CO2). Este oxígeno es utilizado por nuestro organismo para la combustión de lípidos e hidratos de carbono con lo que obtiene energía para alimentar sus células, de este proceso se produce el CO2 que será expulsado difundiéndose desde la circulación venosa hasta los alvéolos pulmonares y hacia el exterior. El nexo de unión entre las células y la atmósfera es la circulación sanguínea, por tanto, existe una coordinación entre los sistemas respiratorio y circulatorio para cubrir de forma adecuada el requerimiento metabólico del organismo en cada circunstancia. A mayor trabajo del organismo mayor es el gasto energético, por tanto, mayor necesidad de transporte de gases.
Además, desempeña otras funciones como la fonación, la regulación del pH y de la temperatura, en que también están implicados otros órganos.
2.1. La función respiratoria
Para llevar a cabo la función respiratoria se diferencian tres procesos de la respiración en su fase pulmonar: la ventilación, la difusión y la circulación.
2.1.1. Ventilación y mecánica ventilatoria
Se denomina ventilación al movimiento del aire a través de las vías de conducción hasta alcanzar al alveolo (inspiración) seguido por el flujo de gases exhalados en dirección opuesta (espiración). Es un proceso que se desarrolla de manera secuencial y cíclica. La ventilación está determinada por el gradiente de presiones existente entre la atmósfera y los alvéolos. En condiciones normales la presión externa o atmosférica se mantiene constante alrededor de 760 mm Hg, considerado el nivel de referencia o presión 0, por tanto para que se dé un flujo de aire la presión que debe modificarse será la presión interna, la presión pulmonar. Cuando existe en el interior del alveolo una presión menor a la presión atmosférica se produce la inspiración, en este caso provocada por la contracción de los músculos inspiratorios, por tanto, es un fenómeno activo. El principal músculo inspiratorio es el diafragma cuya contracción es responsable del 75-80% del movimiento inspiratorio. Al contraerse el diafragma provoca un descenso del suelo de la caja torácica, aumentando así el eje longitudinal de ésta y por tanto su volumen. Los músculos intercostales externos al contraerse elevan la parrilla costal incrementando el volumen de la caja torácica en sentido anteroposterior y transversal.
A continuación, la salida de aire en la espiración es un fenómeno pasivo provocado por la elasticidad pulmonar y la capacidad retráctil de los alvéolos y del surfactante de su interior que hace que actúe la tensión superficial.
La frecuencia respiratoria normal en reposo oscila entre 12-16 ciclos por minuto, con una duración aproximada de 4 segundos por ciclo respiratorio, siendo más larga la espiración (2,5 seg) que la inspiración (1,5 seg).
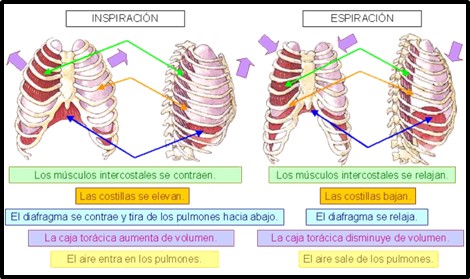
Imagen 6: Mecánica ventilatoria
Fuente: https://www.klipartz.com/es/sticker-png-hxjpr
Hay que tener en cuenta que parte del aire movilizado se utiliza para renovar el aire del espacio que no toma parte activa en el intercambio de gases con la sangre, al que se denomina espacio muerto. Se debe diferenciar entre el espacio muerto anatómico y el espacio muerto funcional. El espacio muerto anatómico corresponde al volumen que ocupa la vía aérea de conducción hasta los bronquiolos. Mientras que el espacio muerto funcional representa todo aquel volumen de aire que no interviene en el recambio gaseoso, es decir, el exceso de aire respecto a su perfusión.
La ventilación alveolar representa el volumen de aire que interviene en la renovación de gases con la sangre, supone la resta entre la ventilación total menos el aire que ocupa el espacio muerto funcional.
Volúmenes y capacidades pulmonares. Espirometría.
El espirograma es el registro del movimiento de volumen de aire que entra y sale de los pulmones que se obtiene mediante una prueba de función pulmonar llamada espirometría.
Para la medición de estos valores se han diferenciado 4 niveles:
1.- Nivel de final de espiración normal
2.- Nivel de final de inspiración normal.
3.- Nivel de inspiración máxima.
4.- Nivel de espiración máxima.
Se denomina volumen a la cantidad de aire comprendida entre dos niveles contiguos y capacidad a la suma de dos o más volúmenes. Se han definido 4 volúmenes y 4 capacidades.
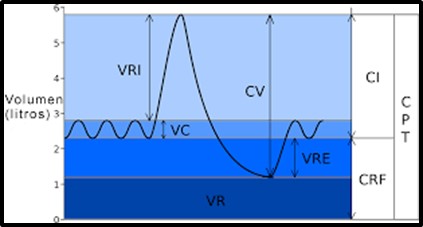
Imagen 7: Volúmenes y capacidades pulmonares.
Fuente: Volums pulmonars est.svg
Volumen corriente (VC): cantidad de aire que ingresa en los pulmones en cada inspiración o que sale en cada espiración, en las condiciones que se especifiquen (reposo, ejercicio). En un varón adulto en reposo comprende, aproximadamente, 500 ml de aire. Esta cantidad de aire representa sólo una décima parte del aire que un pulmón normal puede movilizar, lo que permite recurrir a importantes reservas de inspiración y espiración cuando se aumentan las demandas debido, por ejemplo, al ejercicio físico, fonación, risa, llanto, etc.
Volumen de reserva inspiratoria (VRI): cantidad máxima de aire que se obtiene en una inspiración forzada. Corresponde al aire inspirado adicional al volumen corriente. Alcanza en condiciones normales 3000 ml, aproximadamente
Volumen de reserva espiratoria (VRE): cantidad máxima de aire que se puede expulsar a partir del nivel espiratorio espontáneo normal. Alcanza en condiciones normales 1100 ml, aproximadamente
Volumen residual (VR): Cantidad de aire restante en los pulmones después de una espiración forzada máxima. En promedio alcanza 1200 ml. Este valor no se puede medir directamente con el espirómetro.
Capacidad pulmonar total (CPT): capacidad de aire contenida en el pulmón al final de la inspiración forzada. Corresponde a la suma de los cuatro volúmenes descritos. Aproximadamente 5800 ml.
Capacidad vital (CV): Cantidad total de aire movilizado entre una espiración y una inspiración forzadas. Se obtiene sumando el volumen corriente más el volumen de reserva inspiratoria y espiratoria. Corresponde aproximadamente a 4600 ml. La capacidad vital depende de la talla, edad y sexo del individuo. Obedece a la correcta sincronía entre la generación y la conducción de los estímulos respiratorios, de la capacidad muscular respiratoria, de la mecánica esquelética y del estado del pulmón.
Capacidad inspiratoria (CI): Cantidad máxima de aire que puede inspirarse a partir de una espiración normal. Es la suma del volumen corriente más el volumen de reserva inspiratoria. Su valor aproximado es de 3600 ml.
Capacidad residual funcional (CRF): Volumen de aire que permanece en el pulmón al término de una espiración normal. Representa la suma del volumen de reserva espiratoria más el volumen residual. Corresponde a la posición de reposo del aparato respiratorio, es decir cuando la tendencia de retracción de los pulmones y la tendencia opuesta de la pared torácica a expandirse son iguales. Aproximadamente 2300 ml.
La CRF cumple varias funciones: en la inspiración se diluye el aire fresco inspirado con los aproximadamente 2 l. de aire residual del pulmón, lo que impide que se produzcan cambios bruscos de la composición del aire alveolar manteniendo un equilibrio de los gases de O2 y CO2, y por tanto, manteniendo también un equilibrio ácido-base del organismo. Representa un reservorio de O2, que permite su uso de forma continua también en la espiración o en períodos cortos de apnea. E impide el colapso de los alvéolos pues mantiene el volumen alveolar mínimo para su estabilidad.
Presiones del aparato respiratorio.
En la mecánica respiratoria se diferencian 6 tipos de presiones:
Presión atmosférica: considerada como el punto de referencia 0. El resto de las presiones se expresan como diferencias positivas o negativas en relación a ella.
Presión en la boca o entrada del aparato respiratorio: sin flujo de aire y con la boca abierta corresponde a la presión atmosférica, la de la vía aérea y la de los alvéolos. Dependiendo la fase de la respiración oscila ligeramente por encima o por debajo de la atmosférica.
Presión en las vías aéreas: dependiendo de la dirección del flujo es decreciente hacia el alveolo o hacia la boca.
Presión alveolar: igual a la presión atmosférica en situación estática y con la boca abierta, pero debido, mayormente, a los movimientos del tórax aumentará o disminuirá generando el flujo del aire a través de la vía aérea.
Presión pleural (Ppl): es la presión existente entre las dos hojas de la pleura. Generalmente negativa debido a que el tamaño del pulmón en reposo es menor al del tórax, esto provoca que las propiedades elásticas del pulmón y el tórax traccionan en sentidos opuestos, el pulmón hacia adentro y el tórax hacia afuera.
Presión transpulmonar (Ptp): es la diferencia de presión entre la presión en la boca y la presión pleural. Determina el grado de distensión del pulmón en condiciones estáticas. Durante la respiración debe vencer, además, las resistencias opuestas al movimiento del aire. Corresponde a la diferencia entre la presión alveolar menos la presión pleural.
Resistencias ventilatorias
Durante la mecánica respiratoria los músculos respiratorios deben superar dos tipos de fuerzas que se oponen a ello:
La elasticidad del pulmón y tórax: permite que cuando una estructura se deforma recupere su posición de equilibrio sin requerimiento de gasto energético, en este caso, el tórax y el pulmón recuperan su posición del final de la espiración tranquila (CRF), y por tanto, se opone a la inspiración normal.
La elasticidad de los pulmones se debe a la estructura fibroelástica del parénquima pulmonar, de la tensión superficial en la interfase aire-líquido alveolar, del tejido elástico y conectivo de los vasos y los bronquios y, por último, del contenido de sangre del lecho vascular pulmonar.
Las resistencias friccionales: producidas principalmente por el roce del aire en las vías aéreas (RVA 80%) a la que se suma la producida por la fricción interna de los tejidos del aparato respiratorio (20%). La mayor parte de la resistencia de las vías aéreas se producirá en las vías altas debido a que el flujo de aire en las fosas nasales y la laringe son del tipo turbulento y, por tanto, la resistencia será menor en la respiración por la boca. Debido a que la pared de las vías aéreas está formada por fibras musculares lisas controladas por el sistema nervioso autónomo, la regulación del tono de estas fibras modifica la resistencia y de esta manera el control broncomotor modifica el flujo aéreo (broncoconstricción y broncodilatación).
Por tanto, durante la inspiración los músculos deben vencer la fuerza de retracción elástica y las resistencias friccionales, mientras que la espiración tranquila se producirá por la relajación de los músculos. En la espiración forzada y en ventilaciones superiores a 20 L/min se sumarán los músculos espiratorios teniendo que hacer frente a las mismas resistencias ventilatorias que en la inspiración, pero con fuerza contraria.
Volúmenes dinámicos
La interacción existente entre los volúmenes, las fuerzas y las resistencias respiratorias darán como resultado final el flujo aéreo. Los valores de referencia se relacionan con la edad, peso, talla, peso y grupo étnico. Se denominan dinámicos porque se incluye el factor tiempo, por lo que su medición se realiza en litros/s. Los índices más utilizados en la práctica clínica son:
Capacidad Vital Forzada (CVF): Volumen total que el paciente espira mediante una espiración forzada máxima desde la posición de máxima inspiración.
Volumen Espiratorio Forzado del Primer Segundo (FEV1): Volumen de gas espirado durante el primer segundo de la CVF. Normalmente la espiración forzada total dura entre 5 o 6 segundos, durante el primer segundo se espira aproximadamente un 70-80% de la CV. Es el parámetro más utilizado de función pulmonar. Dado que es dependiente del esfuerzo respiratorio presenta una correlación lineal e inversa a la obstrucción de las vías aéreas.
Índice FEV1/ CVF: Es la relación entre el volumen espirado en el primer segundo con respecto a la capacidad vital forzada. Se conoce con el nombre de Índice de Tiffeneau. Este valor se considera patológico cuando es menor de 0.7. Se identifican 3 patrones clásicos dependiendo de la relación FEV1/ CVF:
- El patrón obstructivo, que produce una limitación en el flujo aéreo durante la espiración, por tanto, disminuirá el FEV1 y la relación FEV1/ CVF.
- El patrón restrictivo, que se produce por la disminución del tamaño del pulmón, de la caja torácica o por enfermedades neuromusculares. Se expresa por una reducción superior al 80% de la CVF teórico.
- El patrón mixto, en el que se producen ambos tipos de alteraciones ventilatorias.
Flujo Espiratorio Forzado entre el 25 y el 75% de la Capacidad Vital Forzada (FEF25-75) o Flujo Máximo de Media Espiración: Flujo espiratorio medio forzado en el intervalo 25-75% de la CVF. Con ello se desecha el primer 25% que es esfuerzo-dependiente y el último 25% que depende del tiempo que el sujeto sostenga el esfuerzo espiratorio, con lo que se centra en la medición del 50% central donde los factores determinantes del flujo máximo son las propiedades mecánicas del pulmón. Es la medida más sensible para detectar la obstrucción precoz de las vías respiratorias pequeñas.
En la Imagen 8 está representado el Bucle formado por la relación de los flujos máximos y los volúmenes dinámicos de una espirometría normal. El estudio de la morfología de la curva Flujo-Volumen permitirá la detección visual de un patrón normal o uno patológico, patrones obstructivos o restrictivos, o detectar obstrucciones intra o extratorácicas.
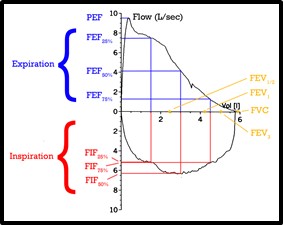
Imagen 8: Espirometría. Bucle de flujo-volúmen. Fuente: Flow_volume_loop.png
2.1.2. La difusión
El proceso de difusión se define como el flujo de partículas de un área de mayor concentración hacia un área de menor concentración.
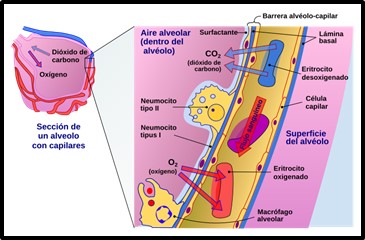
Imagen 9: Proceso de difusión alveolo-capilar. Fuente: Sección transversal de un alvéolo y capilares que muestra la difusión de gases.svg
La difusión pulmonar es el proceso mediante el cual se produce el intercambio de los gases respiratorios entre el alveolo y la sangre a través de la membrana alveolo-capilar.
La capacidad de difusión de los gases respiratorios depende de los siguientes factores:
El espesor y la superficie de la membrana alveolocapilar: De acuerdo con la Ley de difusión de Fick, la transferencia del gas a través de la membrana (VGAS, ml/min) es inversamente proporcional a su espesor (T) y directamente proporcional a la superficie de intercambio (A) en cm2, al gradiente de presiones parciales a cada lado de la membrana (P1-P2, mmHg) y al coeficiente de difusión del gas (D).
VGas = A/T x (P1-P2) x D
El epitelio alveolar, compuesto por la membrana basal y la capa de surfactante, tiene un grosor de entre 0.2-0.3µ, un grosor similar presenta los capilares del intersticio pulmonar, compuestos por una membrana basal y el endotelio. En conjunto la membrana alveolocapilar tiene un espesor de 0.5µ. Esta característica de la estructura pulmonar confiere la máxima eficacia para el intercambio de gases: una amplia superficie y un espesor mínimo de la superficie de intercambio.
El tiempo de contacto del aire alveolar y la sangre: En 0.75 segundos el hematíe atraviesa el capilar en contacto con el alveolo, en tan sólo 0.25 segundos la pO2 y la pCO2 del capilar se igualan a la del alveolo.
La solubilidad del gas en las estructuras que integran la membrana: el O2 difunde mejor en el medio gaseoso (alveolo), en cambio el CO2 es más soluble en el agua y los tejidos por lo que su velocidad de difusión es 20 veces mayor que la del O2. Por este motivo, normalmente los problemas clínicos vienen dados por la afectación en la difusión del O2 y no del CO2.
La diferencia de presión a que se encuentra el gas en el espacio alveolar y en la sangre: en condiciones normales la sangre que llega a los capilares alveolares presenta una presión de oxígeno (pO2) de 40 mmHg y una presión de dióxido de carbono (pCO2) de 45 mmHg, en el aire alveolar la pO2 es de 100 mmHg y una pCO2 de 40 mmHg, debido a la diferencia del gradiente de presiones se producirá la difusión del O2 hacia la sangre y del CO2 hacia el interior del alveolo teniendo como resultado la sangre arterial oxigenada con una pO2 de 100 mmHg y pCO2 de 40 mmHg.
2.1.3. La circulación
La circulación pulmonar es la parte del sistema circulatorio que transporta la sangre desoxigenada desde el ventrículo derecho del corazón por la arteria pulmonar hacia los pulmones y la devuelve oxigenada a la aurícula izquierda a través de las venas pulmonares. Es un sistema de alto flujo y baja presión. El gradiente de presión entre el ventrículo derecho hasta la aurícula izquierda es de 25 mmHg. A nivel de las arterias la presión media es de 15 ± 3 mm Hg (presión sistólica 21± 5, presión diastólica 9 ± 3 mm Hg). En los capilares es de 10 y en las venas de 6 mm Hg. La presión de conducción se establece como la diferencia entre la presión ventricular derecha y la presión auricular izquierda. La resistencia vascular en este circuito es baja. Para movilizar 1 litro de sangre se requieren en el circuito mayor 16 mm Hg, mientras que en el pulmonar son suficientes 2 mm Hg.
El principal factor que regula el flujo sanguíneo pulmonar es la presión parcial de O2 en el gas alveolar (PAO2). Cuando la PAO2 disminuye por debajo del 70% de su valor normal (73 mmHg de PAO2)se produce la vasoconstricción pulmonar.
La vasculatura pulmonar se divide en arterias pulmonares de “conducción” extraparenquimatosa y las arterias pulmonares de “resistencia” intrapulmonar.
Las arterias pulmonares de conducción se dilatan o no se contraen frente a la hipoxia, mientras que las arterias pulmonares de resistencia
Relación ventilación-perfusión
Se llama relación ventilación-perfusión (V/Q) a la relación existente entre la ventilación alveolar y el flujo circulatorio pulmonar (por minuto). Con valores alrededor de 0.8-1 se presume la optimización del intercambio gaseoso en la barrera alveolo-capilar.
Esta relación es desigual en el pulmón debido a la distribución heterogénea del flujo sanguíneo en los pulmones.
El pulmón no es una unidad simple de intercambio gaseoso, sino, por el contrario, unos 300 millones de unidades alveolares con diferentes relaciones V/Q. Por lo tanto, en todas las personas sanas, existe algún disbalance de la V/Q. Esta compleja distribución de ventilación y de perfusión varía ante influencias gravitacionales, cambios en la posición corporal y por cambios en el volumen pulmonar. En pacientes jóvenes normales, la distribución V/Q en el pulmón varía desde 0.6 a 3, y la distribución media está usualmente alrededor de 1. Con la edad, hay un incremento gradual en la desigualdad de V/Q.
Las zonas con baja V/Q pueden contribuir al desarrollo de hipercapnia e hipoxemia, por elevación de la PaCO2 y el descenso de la PaO2. Siempre se producirá hipoxemia, con o sin hipercapnia dependiendo del grado de deterioro y de las características individuales de cada caso.
Como contraste, las unidades con alta V/Q desperdician ventilación, pero tienen escasos efectos adversos en los gases arteriales.El desarrollo de una inadecuada relación V/Q tiene efectos importantes en el intercambio gaseoso pulmonar e interfiere con la transferencia de O2 y del CO 2, produciendo una defectuosa oxigenación, tanto más severa cuanto más acentuado sea el trastorno. Se podría esperar que los pacientes con una inadecuada V/Q tengan hipoxemia e hipercapnia, sin embargo, incluso pequeños incrementos en la PCO2 activan los quimiorreceptores y estimula la ventilación minuta. El incremento de la ventilación minuto es dirigido más a menudo hacia unidades pulmonares que están entre las mejor ventiladas. El incremento de la V/Q en estas unidades conduce hacia un aumento en la PO2 al final del capilar.
En resumen, si se produce un trastorno vascular (Q disminuida) que afecta al pulmón, como una embolia o un infarto pulmonar, diríamos que la relación V/Q aumenta y de modo contrario, si se produce cualquier situación patológica que afecte a la ventilación de los alvéolos, como obstrucción (local, parcial, segmentaria, bronquial) de la vía aérea o incluso la ocupación de alvéolos por líquido (distrés respiratorio, edema agudo de pulmón) diríamos que la relación V/Q disminuye, afectándose en ambos casos el intercambio respiratorio y ocasionando una disminución de oxígeno en la sangre (hipoxemia) acompañada o no de aumento del CO2 (hipercapnia).
Transporte de gases
El objetivo final de la respiración es el suministro de O2 a las células del organismo para que puedan realizar los procesos metabólicos y eliminar el CO2 que resulta del proceso. Los requerimientos del organismo para cubrir sus necesidades en reposo son de 250 ml de O2/minuto.
Por tanto, aunque el transporte de gases no es un proceso puramente propio del sistema respiratorio sino del circulatorio, ambos tienen una relación muy estrecha.
Existen dos formas de transporte de gases en la sangre: disueltos en la sangre (Ley de Henry) o en forma combinada.
El transporte de O2
Una parte del oxígeno difundido desde alveolo a la sangre capilar se disuelve en el plasma en una concentración de 0.3 ml de O2/ 100 ml de sangre. Esta cantidad, aunque es muy pequeña, determina la pO2 en el plasma.
El 98% del transporte de O2 es combinado con la hemoglobina (Hb) unido al hierro de ésta, suponiendo 20 ml de O2/ 100 ml de sangre. Esta forma combinada entre el oxígeno y la hemoglobina se denomina oxihemoglobina (HbO2). Esta combinación es reversible y dependiente de la pO2 en plasma.
La proporción entre el contenido de oxígeno y la máxima capacidad de unión con la Hb es la llamada saturación de la hemoglobina, que en la sangre arterial es del 97% mientras que en la sangre venosa es del 75%.
La relación entre la pO2 y la saturación de la Hb se representa gráficamente en la curva de disociación.
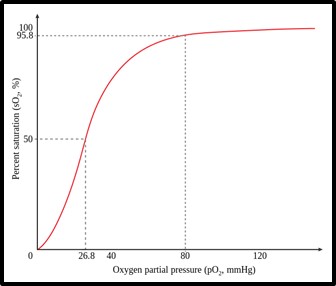
Imagen 10: Curva de disociación de la hemoglobina. Fuente: Hb saturation curve.png
El grado de afinidad de la hemoglobina por el oxígeno puede estimarse a través de un parámetro denominado presión parcial de oxígeno para saturar el 50% de la hemoglobina (P50) situado en 27 mm Hg de pO2.
Los factores más importantes que afectan a la curva de disociación de la hemoglobina son desplazándola hacia la derecha son:
- La presión parcial de CO2 (pCO2). La afinidad del CO2 con la hemoglobina es mayor que la del O2, por lo que si hay un aumento de la concentración de CO2 disminuye la concentración de oxihemoglobina.
- El pH de la sangre. Este factor está ligado al anterior, pues el aumento de pCO2 produce un aumento de H+ (por la acción de la anhidrasa carbónica), que se fijará a la hemoglobina
- El aumento de la temperatura corporal
- La molécula 2,3-difosfoglicerato (2,3-DPG). Esta molécula es un metabolito intermediario de la glucólisis anaerobia del eritrocito que facilita la entrega de O2 a los tejidos. Su concentración es regulable, aumenta con el ejercicio violento prolongado, en la altura y en enfermedades que determinan menos aporte de O2 a las células.
- El monóxido de carbono (CO). Este gas se une a la Hb con una afinidad 210 veces mayor que la del O2, de manera que en concentraciones mínimas de CO en el aire inspirado saturará grandes proporciones de hemoglobina impidiendo de esta manera el transporte de O2. El compuesto formado se denomina carboxihemoglobina.
La alcalosis, la hipocapnia, la hipotermia y/o la disminución del 2,3-DPG tienen el efecto contrario, es decir, desplazan la curva hacia la izquierda, aumentando la afinidad del O2 con la hemoglobina.
El transporte de CO2
El CO2 es producto del metabolismo celular, de las células es transportado hasta el alveolo mediante el torrente sanguíneo, donde se eliminará con la respiración.
El transporte de CO2 se puede dar de las siguientes formas:
- Disuelto en el plasma: al igual que el O2 se solubiliza siguiendo la Ley de Henry en una concentración de 2,9 ml de CO2/100 ml de sangre y dependiente de la concentración del CO2 en plasma y de su coeficiente de solubilidad.
- Forma combinada: un 10% del CO2 es transportado en forma de compuesto carbamínicos en combinación con los grupos amino terminales de las proteínas, mediante una reacción rápida que no requiere catalizador: Hb-NH2 + CO2 = Hb-NHCOO- + H+ (carbaminohemoglobina)
- La mayor parte del CO2 se difunde hacia el interior del hematíe, donde se combina con el agua para producir ácido carbónico, produciéndose la siguiente reacción: CO2+H2O= H2CO3= HCO3- +H+
Una vez formado el bicarbonato las ¾ partes de éste se desplazan al plasma como bicarbonato plasmático mientras que el ión H+ son captados por la hemoglobina.
Como resultado tenemos que, debido al metabolismo celular, en los capilares tisulares se produce un aumento de la PCO2, con la consiguiente disminución del pH facilitando la entrega de O2 (efecto Bohr), a la par que el aumento de Hb reducida facilita la capitación de CO2 (efecto Haldane). En cambio, en los capilares pulmonares donde la PCO2 alveolar (40 mmHg) es menor que la PCO2 de la sangre venosa (46 mmHg) provoca que las reacciones anteriores se den en sentido opuesto y el CO2 se difunde de la sangre hacia el alveolo. A su vez, la hemoglobina reducida se oxigena ayudando a la liberación del CO2 del hematíe. Y, por último, el bicarbonato del plasma entra al eritrocito donde forma H2CO3, que se disocia en CO2 y H2O, que también será eliminado con la respiración.
Equilibrio ácido-base
El equilibrio ácido-base es un fenómeno complejo que implica varios órganos los que mediante su acción fisiológica pretenden mantener constantes una serie de balances interrelacionados, entre ellos el pH, el equilibrio eléctrico, el equilibrio osmótico y la volemia. Si se producen cambios en alguno de éstos se producirán mecanismos compensatorios para devolverlos a límites normales.
El ácido carbónico es el ácido más abundante en el organismo, producido a partir del CO2 proveniente del metabolismo de hidratos de carbono y grasas. Otro ácido importante es el ácido láctico, generado principalmente en los músculos y el hígado. Por último, el metabolismo de proteínas y aminoácidos genera ácidos fosfórico, clorhídrico y sulfúrico.
El enfoque fisiológico tradicional de Henderson y Hasselbalch es el más utilizado en la práctica clínica por ser el más simplificado:
El ion hidrógeno libre (H+) en la sangre arterial se encuentra en una concentración de valores normales entre 35 y 45 mmmol/L, la forma más usada de expresar estas cantidades es mediante el pH, que es el logaritmo negativo de la concentración de H+ en sangre, es decir, en condiciones normales el pH se mantiene entre 7,45 y 7,35.
La concentración de H+ es uno de los determinantes del equilibrio ácido-base, y es dependiente de las interacciones entre la PaCO2, la concentración plasmática de bicarbonato (HCO3-), la disociación constante del ácido carbónico y la solubilidad del CO2.
En condiciones normales, la producción y eliminación de H+ mantiene un equilibrio, por lo que el pH permanece casi constante. En los casos de estrés celular en los que la producción de H+ aumenta, como puede ser durante el ejercicio, se ponen en marcha los mecanismos tampones y la acción reguladora del aparato respiratorio y del riñón.
El sistema regulador del aparato respiratorio se produce a través de la eliminación de CO2 mediante la hiperventilación, o retención de CO2 con la hipoventilación, en respuesta a los cambios detectados por los quimiorreceptores.
El sistema renal utiliza dos mecanismos en la regulación ácido-base: por una parte, tiene la capacidad de reabsorber el bicarbonato filtrado en situaciones de acidemiao excretar el exceso de éste en situaciones de alcalemia. Y, por otro lado, el riñón es capaz de producir nuevas moléculas de bicarbonato a partir de glutamina o a partir de fosfatos.
Aunque se considera que el bicarbonato es el factor principal en el control metabólico (no respiratorio) del equilibrio ácido-base, también es verdad que estas funciones compensatorias son lentas, ya que para alcanzar su máxima eficacia se precisan entre 12 a 72 horas, por tanto, el riñón es un regulador a largo plazo.
BIBLIOGRAFÍA
- Casabona I, Santos R, Lillo M. Historia y evolución de la ventilación mecánica. [Internet]; 2017. Disponible en: https://fcsalud.ua.es/es/portal-de-investigacion/documentos/monografias-libros-y-capitulos-2016/historia-y-evolucion-de-la-ventilacion-mecanica.pdf.
- Dräger. https://www.draeger.com/Library/Content/technology-for-life-br-9070355-en.pdf. [Online].
- García-Talavera, I., Lobato, S. D., Bolado, P. R., Villasante, C. Músculos respiratorios. Arch Bronconeumol [Internet]. 1992; 28(5), 239-246. Disponible en: https://www.sciencedirect.com/science/article/abs/pii/S0300289615313351
- Mena, E. C., & Bolton, R. M. Aparato respiratorio. Mediterráneo [Internet] 1999; 27, 185-189. Disponible en: https://d1wqtxts1xzle7.cloudfront.net/36915107/Aparato_Respiratorio_Fisiologia_y_Clinica_5%C2%AA_Ed.pdf?1425936221=&response-content-disposition=inline%3B+filename%3DAparato_Respiratorio_Fisiologia_y_Clinic.
- Turchetto E.S, Tusman G. Respiratorio. En: Ceraso D.H. Terapia Intensiva. 4º Edición. Buenos Aires. Médica Paramericana; 2007. p.175-359.
- Clemente F.J. Iniciación a la Ventilación Mecánica: Aplicaciones y Cuidados. 2º Edición. Formación Continuada Logoss; 2006.
- Sánchez T., Concha I. Estructura y Funciones del sistema respiratorio. Neumol Pediatr [Internet]. 2018; 13 (3): p. 101-106. Disponible en: https://neumologia-pediatrica.cl/index.php/NP/article/view/212
- García E., Sandoval J.C., Díaz L.A., Salgado J.C. Ventilación mecánica invasiva en EPOC y asma. Med Intensiva [Internet].2011; 35(5): p.288-298. Disponible en: https://medintensiva.org/es-ventilacion-mecanica-invasiva-epoc-asma-articulo-S0210569110002871
- Goñi R., Yoldi E., Casajús L., et al. Fisioterapia respiratoria en la unidad de cuidados intensivos: Revisión bibliográfica. Enferm Intensiva [Internet]. 2018; 29 (4): p.169-181. Disponible en: https://www-sciencedirect-com.sabidi.urv.cat/science/article/pii/S1130239918300580
- Colaianni N., Castro M. Terapia nasal de alto flujo en la insuficiencia respiratoria aguda hipoxémica por SARS-CoV-2. Rev Patol Respir [Internet]. 2020; 23 (3): p. 99-107. Disponible en: https://www.revistadepatologiarespiratoria.org/descargas/PR_23-3_99-107.pdf
- Guérin C., Reignier J., Richard JC., et al. Posicionamiento en decúbito prono en el síndrome de dificultad respiratoria aguda grave. N Engl J Med [Internet]. 2013; 368: p.2159-2168. Disponible en: https:// www.nejm.org/doi/full/10.1056/nejmoa1214103
- Thompson B.T., Chambers R.C., Liu K.D. Síndrome de distrés respiratorio agudo. N Engl J Med [Internet]. 2017; 377: p. 562-572. Disponible en: https://www.nejm-org.sabidi.urv.cat/doi/10.1056/NEJMra1608077?uri_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub0pubmed
- Colaboradores de Wikipedia. Relación ventilación-perfusión [Internet]. Wikipedia, La enciclopedia libre, 2021. Disponible en: https://es.wikipedia.org/w/index.php?title=Relaci%C3%B3n_ventilaci%C3%B3n-perfusi%C3%B3n&oldid=134126240.
- Colaboradores de Wikipedia. Sistema de control de la respiración [Internet]. Wikipedia, La enciclopedia libre; 2021 feb 18, 12:52 UTC [cited 2021 abr 15]. Disponibleen: https://es.wikipedia.org/w/index.php?title=Sistema_de_control_de_la_respiraci%C3%B3n&oldid=133321088.
- Aristizábal-Salazar, R. E., Calvo-Torres, L. F., Valencia-Arango, L. A., Montoya-Cañon, M., Barbosa-Gantiva, O., Hincapié-Baena, V. Equilibrio ácido-base: el mejor enfoque clínico. Revista Colombiana de Anestesiología, [Internet]; 2015 43(3), 219-224. Disponible en:https://www.sciencedirect.com/science/article/pii/S0120334715000441
- Cruz Mena E, Moreno B. Aparato Respiratorio: Fisiología y Clínica, cuarta edición. [Internet]Publicación Técnica Mediterráneo, Santiago de Chile. 1999. Disponible en:http://www.untumbes.edu.pe/vcs/biblioteca/document/varioslibros/0562.%20Aparato%20respiratorio.%20Fiaiolog%C3%ADa%20y%20cl%C3%ADnica.pdf
- Colaboradores de Wikipedia. Hipoxia. Wikipedia, La enciclopedia libre [Internet]; 2021 [fecha de consulta: 16 de abril del 2021]. Disponible en https://es.wikipedia.org/w/index.php?title=Hipoxia&oldid=133783721.
- Colaboradores de Wikipedia. Hipercapnia. Wikipedia, La enciclopedia libre [Internet] 2020. Disponible en https://es.wikipedia.org/w/index.php?title=Hipercapnia&oldid=131748438.
- Masclans, J. R., Pérez-Terán, P., Roca, O. Papel de la oxigenoterapia de alto flujo en la insuficiencia respiratoria aguda. Medicina Intensiva [Internet] 2015; 39(8), 505-515. https://www.sciencedirect.com/science/article/abs/pii/S0210569115001217
- Arraiza Gulina, N. Guía rápida y póster de dispositivos de oxigenoterapia para enfermería. [Internet] 2015. Disponible en: https://academica-e.unavarra.es/handle/2454/18478
- Palacio Herrero, C. Intervención de la enfermería en la cámara hiperbárica.2015. Disponible en: https://repositorio.unican.es/xmlui/handle/10902/7470
- Vásquez Teran A, Díaz Rodríguez S, Alarcón Hinojosa M, Iza Niza A, Herrera Bastidas G, Casa Tipán J. Cuidados de Enfermería al inicio de Ventilación Mecánica Invasiva en la Unidad de Cuidados Intensivos. CAMbios-HECAM [Internet]. 14may2020 [citado 24abr.2021]; 18(2):106-15. Disponible en: https://revistahcam.iess.gob.ec/index.php/cambios/article/view/558
- Aristizábal-Salazar, R. E., Calvo-Torres, L. F., Valencia-Arango, L. A., Montoya-Cañon, M., Barbosa-Gantiva, O., Hincapié-Baena, V. Equilibrio ácido-base: el mejor enfoque clínico. Rev Colomb Anestesiol. [Internet] 2015; 43(3), 219-224. Disponible en: https://www.sciencedirect.com/science/article/pii/S0120334715000441
- Mugarra, G. Dispositivos para la ventilación de urgencia no quirúrgica. Manejo de la vía aérea. Disponible en: https://sofos.scsalud.es/sofosdocs/Urgencias/MVAA2015_5/modulos/3/MVAA%20M3%20T1.pdf
- Elsevier Connect. Inserción de la mascarilla laríngea de intubación Fastrach. (MLF). 2017. Disponible en: https://www.elsevier.com/es-es/connect/medicina/insercion-de-la-mascarilla-laringea-de-intubacion-fastrach-mlf
- Colaboradores de Wikipedia. Intubación traqueal. Wikipedia, la enciclopedia libre. 23 de abril de 2021. Disponible en: https://en.wikipedia.org/w/index.php?title=Tracheal_intubation&oldid=1019436227 .
- Melgar, A. B., Galván, M. J., Gandullo, E. V., Hidalgo, A. G. Manejo del paciente traqueostomizado, cánulas y aplicación de fármacos inhalados. Manual de diagnóstico y terapéutica en neumología. 3º edición. Neumosur, [Internet] 2016; 293-299. Disponible en:https://www.neumosur.net/files/publicaciones/ebook/25-TRAQUEOSTOMIZADO-Neumologia-3_ed.pdf
- Gutiérrez Muñoz Fernando R.. Insuficiencia respiratoria aguda. Acta méd. peruana [Internet]. 2010 Oct; 27(4 ): 286-297. Disponible en:
- http://www.scielo.org.pe/scielo.php?script=sci_arttext&pid=S1728-59172010000400013&lng=es.
- Hernández-López GD, Zamora GSE, Gorordo SLA, et al. Síndrome de insuficiencia respiratoria aguda. Rev Hosp Jua Mex. [Internet] 2015;82(1). Disponible en: https://www.medigraphic.com/cgi-bin/new/resumen.cgi?IDARTICULO=57596
- Carpio, C., D. Romera, J. Fernández-Bujarrabal. "Insuficiencia respiratoria aguda." Medicine-Programa de Formación Médica Continuada Acreditado 10.63. [Internet] 2010; 4332-4338. Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=3316690
- Palencia-Herrejón, E., Borrallo-Pérez, J., Pardo-Rey, C., & de la SEMICYUC, G. D. Intubación del enfermo crítico. Med Intensiva. [Internet] 2005; 32(Supl. 1), 3-11. Disponible en: https://www.medintensiva.org/es-pdf-13116121
- Rooco, C., Guzmán, J. Manejo de la vía aérea por acceso infraglótico: ventilación jet y cricotirotomía. Rev Chil Anest [Internet] 2010; 39, 158-66. Disponible en: http://www.sachile.cl/upfiles/revistas/4ce1456608bc9_rocco.pdf
- Moya, C. Sistemas de humidificación en ventilación mecánica. Mirada de un terapeuta respiratorio. Teoría y praxis investigativa, 2008; 3(2), 73-82. Disponible en: https://Dialnet-SistemasDeHumidificacionEnVentilacionMecanicaMirad-3701012.pdf
- Quintana Carrasco, J. K. Manejo fisioterapéutico de humidificación en ventilación mecánica invasiva, 2018. Disponible en: https://core.ac.uk/download/pdf/230598397.pdf
- Dávila Cabo de Villa Evangelina, López González Rosendo, Márquez Ercia Fernando, Hernández Dávila Carlos. Intubación de secuencia rápida. Medisur [Internet]. 2015 Ago ; 13( 4 ): 533-540. Disponible en: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S1727-897X2015000400010&lng=es.
- González, M. G. Procedimiento de traqueostomía percutánea: control y seguimiento de enfermería. Enfermería intensiva [Internet]. 2009; 20(2), 69-75. Disponible en: https://www.sciencedirect.com/science/article/abs/pii/S1130239909711484
- Hernández, C. P., Santos, M. M., Huelva, A. B. Traqueostomía: Indicaciones, técnica y complicaciones. Intubación. Disponible en: https://seorl.net/PDF/Laringe%20arbor%20traqueo-bronquial/110%20-%20TRAQUEOTOM%C3%8DA%20INDICACIONES,%20T%C3%89CNICA%20Y%20COMPLICACIONES.%20INTUBACI%C3%93N.pdf
- Melgar, A. B., Galván, M. J., Gandullo, E. V., Hidalgo, A. G. Manejo del paciente traqueostomizado, cánulas y aplicación de fármacos inhalados. Manual de diagnóstico y terapéutica en enumología. 3º edición. Neumosur [Internet] 2016; 293-299. Disponible en: https://www.neumosur.net/files/publicaciones/ebook/25-TRAQUEOSTOMIZADO-Neumologia-3_ed.pdf
- Por I., Ramchandani A., Moreno M. Ventilación Mecánica: Conocimientos básicos. https://especialidades.sld.cu/enfermeriaintensiva/files/2014/04/vent_mecanic_princ_basic.pdf
- Gutiérrez Muñoz Fernando. Ventilación mecánica. Acta méd. peruana [Internet]. 2011 Abr; 28 (2): 87-104. Disponible en: http://www.scielo.org.pe/scielo.php?script=sci_arttext&pid=S1728-59172011000200006&lng=es.
- Sánchez, J. I., Lozano, R. M., Tortosa, F. C., Morales, M. R., Dormal, F. P., & Amorós, A. L. PROBLEMAS PSICOLÓGICOS EN PACIENTES SOMETIDOS A VENTILACIÓN MECÁNICA. Enfermería Global [Internet] 2002; 1. Disponible en: https://digitum.um.es/digitum/bitstream/10201/23857/1/680-3168-3-PB.pdf
- Hernández-López GD, Zamora GSE, Gorordo SLA, et al. Síndrome de insuficiencia respiratoria aguda. Rev Hosp Jua Mex. 2015;82(1). Disponible en: https://www.medigraphic.com/cgi-bin/new/resumen.cgi?IDARTICULO=57596
- Rodriguez-Buenahora Ruben Dario, Ordoñez-Sánchez Sergio Alexander, Gómez-Olaya Jimmy Leandro, Camargo-Lozada Marly Esperanza. Decúbito prono en el Síndrome de Dificultad Respiratoria Aguda, de la fisiología a la práctica clínica. Medicas UIS [Internet]. 2016 Aug; 29 (2): 81-101. Disponible en: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0121-03192016000200009&lng=en.
- Martinón-Torres, F., Martinón-Sánchez, J. M. Toracocentesis y drenaje pleural. An Pediatr Contin [Internet] 2003; 1(3), 159-165. Disponible en: https://www.elsevier.es/es-revista-anales-pediatria-continuada-51-articulo-toracocentesis-drenaje-pleural-S1696281803716084
- Estrada Masllorens, J. M., Galimany Masclans, J., Sarria Guerrero, J. A. Drenaje torácico cerrado. Sistema de recogida no reutilizable: Pleur-evac®. Nursing [Internet] 2012; vol. 30, num. 6, p. 54-58. Disponible en: http://diposit.ub.edu/dspace/handle/2445/33264
- Iñiguez, F. Terapia inhalatoria en los pacientes con ventilación mecánica domiciliaria. Neumol Pediátr, [Internet] 2012; 87. Disponible en: https://www.researchgate.net/profile/Fernando-Iniguez/publication/308715130_Terapia_inhalatoria_en_los_pacientes_con_Ventilacion_Mecanica_Domiciliaria/links/57ec76b108ae92eb4d265ba5/Terapia-inhalatoria-en-los-pacientes-con-Ventilacion-Mecanica-Domiciliaria.pdf
- Retuerto Platero, C. Guía de cuidados de enfermería al paciente adulto en terapia de oxigenación por membrana extracorpórea (ECMO) 2016. Disponible en: https://core.ac.uk/download/pdf/211101545.pdf
- Lema, B. D., Riu, P., Casan, P. Tomografía por impedancia eléctrica. Estandarización del procedimiento para su aplicación en neumología. Arch. Bronconeumol. [Internet] 2006; 42(6), 299-301. Disponible en: https://medes.com/publication/29610
- Riera J., Riu P.J., Casan P., Masclans J.R. Tomografía de impedancia eléctrica en la lesión pulmonar aguda. Med. Intensiva [Internet]. 2011 nov; 35(8): 509-517. Disponible en: http://scielo.isciii.es/scielo.php?script=sci_arttext&pid=S0210-56912011000800009&lng=es.
- Monsalve-Naharro José Ángel, Domingo-Chiva Esther, García Castillo Sergio, Cuesta-Montero Pablo, Jiménez-Vizuete José María. Óxido nítrico inhalado en pacientes adultos con síndrome de distrés respiratorio agudo. Farm Hosp. [Internet]. 2017 Abr; 41( 2 ): 292-312. Disponible en: http://scielo.isciii.es/scielo.php?script=sci_arttext&pid=S1130-63432017000200292&lng=es.
- López, M. R., Canzobre, S. R. Ferreiro, A. P. Protocolo de manejo del paciente con terapia de óxido nítrico. Enfermería en cardiología: revista científica e informativa de la Asociación Española de Enfermería en Cardiología, [Internet] 2017; (72), 56-62. Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=6308145
- López, L. R. El decúbito prono en el Síndrome de Distrés Respiratorio del Adulto: cuidados de enfermería. Enferm Intensiva [Internet] 2002; 13(4), 146-154. Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=636868
- Iñiguez, F. Terapia inhalatoria en los pacientes con ventilación mecánica domiciliaria. Neumol Pediátr. 2012; 87. Disponible en: https://www.researchgate.net/profile/Fernando-Iniguez/publication/308715130_Terapia_inhalatoria_en_los_pacientes_con_Ventilacion_Mecanica_Domiciliaria/links/57ec76b108ae92eb4d265ba5/Terapia-inhalatoria-en-los-pacientes-con-Ventilacion-Mecanica-Domiciliaria.pdf
- Estrada Masllorens, J. M., Falcó Pegueroles, A. M., Moreno Arroyo, M. C. Drenaje torácico cerrado. Procedimiento y cuidados de enfermería a propósito de un caso. Nursing [Internet] 2012, vol. 30, num. 5, p. 50-57. Disponible en: https://core.ac.uk/download/pdf/16208296.pdf